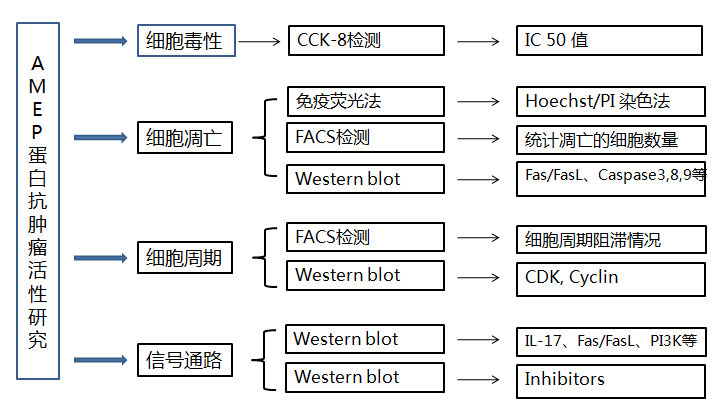
抗菌肽(antimicrobial peptides, AMPs)是存在于生物体内具有抗菌活性的小分子多肽的总称[1]。是生物体内先天免疫防御机制的重要组成部分,当病原菌侵入机体时,抗菌肽能够直接作用于病原菌,抑制其增殖甚至能够直接将其杀死[2]。目前,已从细菌、昆虫、植物和脊椎动物,包括人类中分离获得3000余种抗菌肽,这类活性多肽多数由12~100个氨基酸残基组成[3],大部分带正电荷,含有一个富含赖氨酸、精氨酸等阳离子的N端,C端则由非极性氨基酸丙氨酸、甘氨酸等构成,且 C 端大部分酰胺化[4]。抗菌肽具有强碱性(pI 8.9-10.7)、热稳定性、酸碱适应性、广谱抗菌性以及不易产生耐药性等特点。通过对抗菌肽的深入研究发现,抗菌肽不仅具有抗菌活性[5],某些抗菌肽还能激活免疫系统,参与细胞信号传导,调节细胞增殖,抗病毒,抗寄生虫,及抗肿瘤[6-10]等功能。
课题组成员长期进行抗菌肽的分离、鉴定、活性分析及功能性产品的研制与开发。2019年课题组在枯草芽孢杆菌中发现了一种具有较强抗菌活性的蛋白组分,单一蛋白组分经质谱测序和数据库比对分析,并未发现与该蛋白高度相似的抗菌序列,即该蛋白为一个功能未被定义的全新蛋白,将其命名为AMEP,GenBank登录号为WP_017418614.1。AMEP由76个氨基酸组成,分子量为8.36 kD,等电点为10.05,富含赖氨酸和亮氨酸等疏水氨基酸,其中Lys为12个占氨基酸总数的15.8%,17-36位氨基酸被预测为跨膜结构域,分子式为C402H629N95O94S2,原子总数为1222个,在大肠杆菌中的半衰期大于10小时,蛋白质亲水性的平均值为0.359。不稳定指数为1.35,为稳定的蛋白质。AMEP的二级结构主要由α螺旋组成,以多聚体形式稳定存在于水性溶液中,且具有良好的热稳定性。
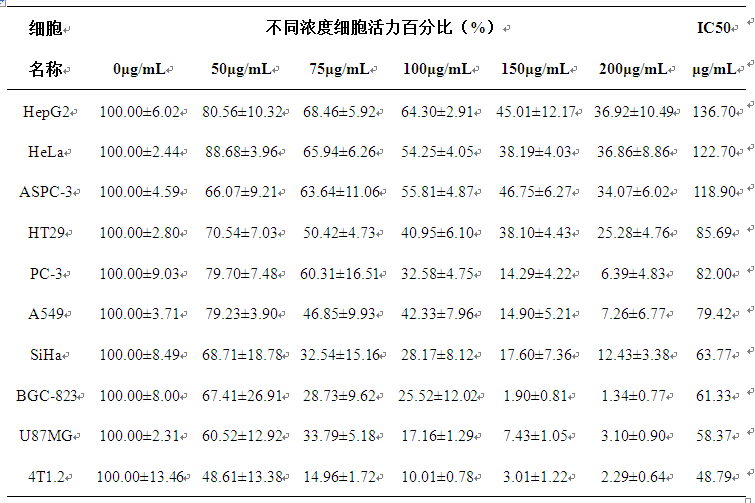
研究显示,大多数具有抗癌活性的抗菌肽均具有α-螺旋的构象[11]。如Aurein家族抗癌肽具有两亲的α-螺旋结构,具有光谱的抗癌活性,对人体中 90%的癌细胞具有杀伤作用。如天蚕素 B1 (CB1) 具有极性脂质的表面,其活性主要来自于 cecropin B(CB)两个 α 螺旋的肽段,增加的 α 螺旋结构可增强 α-螺旋肽的稳定性,促使其更灵活有效地插入质膜,导致膜溶解,从而杀伤肿瘤细胞[12-13]。由 Bcl-2 蛋白家族促凋亡成员之一 BIM 与 Bcl-2 稳定 α 螺旋结构域 (SAHBs)组成的 BIM-SAHB A 结构域可靶向Bcl-2 通路,通过沉默血液癌症表达的抗凋亡基因,导致肿瘤细胞死亡[14-15]。另有研究报道[16]富含亮氨酸和赖氨酸的抗菌肽可抑制肿瘤细胞的转移。基于以上研究成果,我们推测,具有α-螺旋结构和富含亮氨酸/赖氨酸的AMEP蛋白也可能具有抗肿瘤活性。课题组采用不同浓度的AMEP蛋白处理肿瘤细胞,结果显示AMEP对所试的十种肿瘤细胞的增殖均具有不同程度的抑制作用(见工作基础),说明AMEP蛋白确实具有抗肿瘤活性。
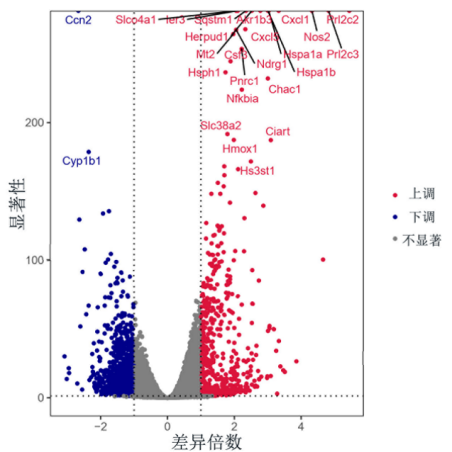
与杀菌机制相比,昆虫抗菌肽抑制和杀死肿瘤细胞的机制要复杂得多。一是特异性破坏肿瘤细胞膜结构[17]。抗菌肽携带大量的正电荷[18]能够与携带大量负电荷的肿瘤细胞膜相结合,在肿瘤细胞膜上形成孔洞[19]破坏细胞膜的完整性从而杀死肿瘤细胞。二是抑制肿瘤细胞增殖。抗菌肽可以穿过肿瘤细胞的细胞膜进入细胞内部,作用于多个靶位点来破坏肿瘤细胞的内部结构,并且能够诱导核染色体 DNA 的断裂,以此来抑制肿瘤细胞 DNA 的合成,从而影响肿瘤细胞的增殖[20]一些抗菌肽能够通过抑制肿瘤血管的生成来降低肿瘤细胞的迁移和侵袭能力三是诱导肿瘤细胞凋亡。研究表明某些抗菌肽可以进入肿瘤细胞内部激活细胞内凋亡信号通路,引起细胞程序性死亡。那么,AMEP蛋白是通过何种途径发挥抗肿瘤活性的?课题组还进一步针对抑制效果突出的肿瘤细胞进行了转录组学分析,结果显示,AMEP可以激活肿瘤细胞凋亡或坏死相关的信号通路(见工作基础)。
综合国内外的文献报道及我们的前期研究结果,我们推测AMEP可抑制多种肿瘤细胞的增殖,并且通过诱导肿瘤细胞的凋亡从而发挥了抗肿瘤作用。然而,AMEP蛋白具体的抗肿瘤机制还有待于深入研究。本项目旨在探讨AMEP蛋白的抗肿瘤活性,并初步揭示其抗肿瘤机制,为拓展AMEP蛋白的应用提供实验基础。
参考文献:
[1] Harris F, Dennison SR, Singh J, et al. On the selectivity and efficacy of defense peptides with respect to cancer cells. Med Res Rev, 2011, 33(1): 190-234
[2] Riedl S, Rinner B, Asslaber M, et al. In search of a novel target-phosphatidylserine exposed by non-apoptotic tumor cells and metastases of malignancies with poor treatment efficacy. Biochim Biophys Acta, 2011, 1808(1): 2638-45
[3] Al-Benna S, Shai Y, Jacobsen F, et al. Oncolytic activities of host defense peptides. Int J Mol Sci, 2011, 12(11): 8027-51
[4] Hammami R, Fliss I. Current trends in antimicrobial agent research: chemo- and bioinformatics approaches. Drug Discov Today, 2010, 15(13-14): 540-6
[5] Hoskin DW, Ramamoorthy A. Studies on anticancer activities of antimicrobial peptides. Biochim Biophys Acta, 2008, 1778(2): 357-75
[6] Schweizer F. Cationic amphiphilic peptides with cancer�selective toxicity. Eur J Pharmacol, 2009, 625(1-3): 190-4
[7] Dobrzynska I, Szachowicz-Petelska B, Sulkowski S, et al. Changes in electric charge and phospholipids composition in human colorectal cancer cells. Mol Cell Biochem, 2005, 276(1-2): 113-9
[8] Papo N, Seger D, Makovitzki A, et al. Inhibition of tumor growth and elimination of multiple metastases in human prostate and breast xenografts by systemic inoculation of a host defense-like lytic peptide. Cancer Res, 2006, 66(10): 5371-8
[9] Al-Hajj M, Clarke MF. Self-renewal and solid tumor stem cells. Oncogene, 2004, 23(43): 7274-82
[10] Gross S, Andra J. Anticancer peptide NK-2 targets cell surface sulphated glycans rather than sialic acids. Biol Chem, 2012, 393(8): 817-27
[11] Papo N, Braunstein A, Eshhar Z, et al. Suppression of human prostate tumor growth in mice by a cytolytic D-, L-amino acid peptide: membrane lysis, increased necrosis, and inhibition of prostate-specific antigen secretion. Cancer Res, 2004, 64(16): 5779-86
[12] Alvarez-Calderon F, Gregory MA, Degregori J. Using functional genomics to overcome therapeutic resistance in hematological malignancies. Immunol Res, 2013, 55(1-3): 100-15
[13] Schroder-Borm H, Bakalova R, Andra J. The NK-lysin derived peptide NK-2 preferentially kills cancer cells with increased surface levels of negatively charged phosphati�dylserine. FEBS Lett, 2005, 579(27): 6128-34
[14] Lemeshko VV. Electrical potentiation of the membrane permeabilization by new peptides with anticancer properties. Biochim Biophys Acta, 2013, 1828(3): 1047- 56
[15] Wu JM, Jan PS, Yu HC, et al. Structure and function of a custom anticancer peptide, CB1a. Peptides, 2009, 30(5): 839-48
[16] Srisailam S, Arunkumar AI, Wang W, et al. Conformational study of a custom antibacterial peptide cecropin B1: implications of the lytic activity. Biochim Biophys Acta, 2000, 1479(1-2): 275-85
[17] Edison N, Reingewertz TH, Gottfried Y, et al. Peptides mimicking the unique ARTS-XIAP binding site promote apoptotic cell death in cultured cancer cells. Clin Cancer Res, 2012, 18(9): 2569-78
[18] Labelle JL, Katz SG, Bird GH, et al. A stapled BIM peptide overcomes apoptotic resistance in hematologic cancers. J Clin Invest, 2012, 122(6): 2018-31
[19] Iwasaki T, Ishibashi J, Tanaka H, et al. Selective cancer cell cytotoxicity of enantiomeric 9-mer peptides derived from beetle defensins depends on negatively charged phosphatidylserine on the cell surface. Peptides, 2009, 30(4): 660-8